
Введение. Исследования по генетике популяций народов Кавказа пока являются единичными. Изучение частот групп крови в кавказских популяциях началось давно (1). Полиморфные локусы белков и ферментов крови пока исследованы лишь у нескольких народов Кавказа и Закавказья (2, 3, 4). Наибольшее число локусов было изучено нашими бывшими сотрудниками из группы биохимической генетики лаборатории генетики человека Института общей генетики по программе, разработанной нами при исследовании популяций народов Сибири (5, 6), у грузин, армян, азербайджанцев, адыгейцев, карачаевцев, черкесов, чеченцев, абазинов, балкарцев, кабардинцев (7, 8, 9, 10).
Мы исследовали полиморфизм нескольких локусов белков крови у талышей Юго-Востока Азербайджана. У населения Грузии были исследованы антропологические признаки. Мы вычислили генетические расстояния популяций народов Кавказа и Закавказья между собой и рядом соседних этносов. По матрице генетических расстояний мы построили эволюционную дендрограмму народов Кавказа по 28 аллелям 12 локусов белков, ферментов и групп крови. Далее по матрице генетических расстояний 55 человеческих популяций, принадлежащих к четырем большим человеческим расам, вычисленной нами ранее (11) мы построили эволюционную дендрограмму этих популяций. В целом кластеризация популяций Кавказа в этих двух дендрограммах не противоречит представлениям отечественных антропологов. В статье развивается высказанная одним из авторов этой статьи А.Ф. Назаровой в 1999 г. гипотеза (12) о палеоазиатском происхождении европеоидов (кавказоидов). Кластеризация других европеоидных популяций в целом, соответствует общепринятым представлениям, в частности, кластеризация славянских популяций подтверждает антропологические результаты Т.И. Алексеевой (13, 14).
Материалы и методы. Популяция талышей села Пирасора Лерикского района Азербайджанской ССР изучалась одним из авторов данной работы А.Ф. Назаровой во время экспедиции лаборатории генетики Института проблем экологии и эволюции РАН летом 1987 г., руководитель экспедиции — д.б.н. Ф.А. Ата-Мурадова. Пробы крови из вены по 7-10 мл центрифугировали для отделения плазмы крови от эритроцитов, и сразу замораживали. Электрофорез белков производила по возвращении в Москву А.Ф. Назарова. Электрофорез в вертикальном блоке полиакриламидного геля в соответствии с методом Ганэ с соавт.(15) с некоторыми модификациями.
Гаптоглобин окрашивали раствором, содержащим 1 мл диметилформамида, 50 мг 3-амино-9-этил-4-карбазола и 1 мл 30% H O в 30 мл 10% CH COOH. Группо-специфический компонент (Gc), трансферрин (Tf) и альбумин(Alb) окрашивали с помощью кумасси. Популяционно- генетические вычисления частот генов, относительной и средней гетерозиготности талышей и их генетические расстояния до других популяций проводила А.Ф. Назарова. Среднюю гетерозиготность рассчитывали по методу (16). Относительную гетерозиготность вычисляли по методу (17), генетические расстояния вычисляли по Кавалли-Сфорца и Бодмеру (18). Дендрограммы популяций по матрицам генетических расстояний в соответствии с методом попарной кластеризации, примененным Машуровым и Черкащенко в (19) строили А.Ф. Назарова и С.М. Алхутов. Краниометрическое исследование населения Грузии выполнял В.О. Асланишвили.
Результаты исследований и их обсуждение. Популяция талышей обитает на Юго-Востоке Азербайджана и на Севере Ирана. Талыши говорят на варианте фарси-талышском языке, принадлежащем к северо-западной группе иранских языков. В Азербайджане все талыши владеют и азербайджанским языком. Талыши являются мусульманами-шиитами. В переписи населения, проводившейся в СССР в 1970 г., талыши не выделены в особую этническую группу, и в нынешнем независимом Азербайджане талыши также считаются азербайджанцами. Однако и их принадлежность к индо-европейской языковой семье, и антропологические признаки (большая европеоидность, чем у азербайджанцев), и многие элементы материальной культуры заставляют считать талышей отдельным, хотя и немногочисленным этносом. Объем популяции талышей в целом превышает 200 000 человек, в том числе на Севере Ирана около 70 000 чел. Традиционные занятия талышей- скотоводство (разведение овец, коров) и земледелие в долинах, в том числе садоводство. Талыши Азербайджана живут в горных селениях, иногда на высоте 2000 м и более над уровнем моря, на плато Талышских гор и в долинах, в районном центре Лерик, в городах Ленкорань и Астара. Среди талышей распространено долгожительство, чему, возможно, способствует природная среда обитания горцев, умеренное питание, в основном растительно-молочное, постоянный физический труд. Однако, повидимому, существует и генетический компонент долгожительства талышей, поскольку в отдельных семьях долгожительство повторяется в поколениях.
Мы исследовали популяции талышей в генетическом аспекте впервые, краткое сообщение было опубликовано нами ранее (20). Потом во время следующей экспедиции нашей лаборатории были исследованы демографические аспекты популяции талышей (21). Распределение частот генов белков крови в популяции талышей высокогорного села Пирасора (1800 м над уровнем моря) представлено в Табл. 1. Объем исследованной выборки — 40 человек.
Нарушений равновесия Харди-Вайнберга, как нами показано ранее, в популяции талышей Пирасоры нет (20). Частота аллеля НР 1 соответствует таковой у европеоидов, особенно у жителей Ирана и Афганистана (Табл1, 22). В выборке талышей Пирасоры мы не обнаружили гомозигот 2-2 по локусу GC, но, поскольку число гетерозигот 1-2 также было не велико, популяционное равновесие не нарушено (20). Трансферрин в изученной выборке оказался мономорфным. Не встретили мы у талышей и редких вариантов альбумина.
Популяция талышей Пирасоры изолирована в связи со своим географическим положением — она находится в 20 км по горной дороге от районного центра Лерик. Эта популяция в значительной мере обладает сниженной гетерозиготностью вследствие традиционно эндогамных браков. Кузенные браки в Пирасоре — обычное явление.
Таблица 1.
|
Частоты генов популяции талышей Пирасоры |
||
|
Локус |
Аллель |
Частота аллеля |
|
HP |
HP1 |
0,4375 |
|
HP2 |
0,5625 |
|
|
GC |
GC1 |
0,8875 |
|
GC2 |
0,1125 |
|
|
TF |
TFc |
1,0000 |
|
TFd |
0,0000 |
|
|
TFb |
0,0000 |
|
|
Alb |
Alba |
1,0000 |
|
Albредк |
0,0000 |
|
Мы вычислили относительную гетерозиготность популяции Пирасоры по локусам HP и GC. Как видно из Табл. 2, наблюдаемая гетерозиготность популяции талышей Пирасоры ниже теоретической. Относительная гетерозиготность по локусу GC равна -0,3073.
Таблица 2.
Относительная гетерозиготность популяции талышей Пирасоры
|
Локус |
ho |
ht |
D |
|
HP |
0,175 |
0,1914 |
0,0666 |
|
GC |
0,225 |
0,3248 |
-0,3073 |
В табл. 3 приведена вычисленная нами средняя гетерозиготность популяции Пирасоры по всем изученным локусам.
Таблица3.
Средняя гетерозиготность популяции талышей Пирасоры
|
Локус |
H для отдельного локуса |
Н средняя на локус |
|
HP |
0,492188 |
|
|
GC |
0,1996875 |
|
|
TF |
0,0 |
|
|
Alb |
0,0 |
Средняя гетерозиготность по всем локусам равна 0,17297 — очень низкая, что свидетельствует о значительной гомозиготности талышей Пирасоры. Гомозиготность, повидимому, вызвана изолированностью популяции Пирасоры и традиционной склонностью к инбридингу. В целом Пирасору можно считать изолятом.
Как известно, в середине XVIII века на Юго-Западном побережье Каспийского моря в связи с распадом государства Надир-Шаха образовалось талышское ханство. В 1795 г. талышский хан Мир-Мустафа обратился к России с просьбой о покровительстве, в связи с чем Талышское ханство в 1802 г. было принято под протекторат, а в 1828 г. по Туркманчайскому договору было присоединено к России. Однако сохранились как культурные, так и социально-экономические связи популяции талышей, живущей в России, с талышами Ирана, не прекращались и взаимные миграции. Поэтому мы решили вычислить генетические расстояния популяции талышей Пирасоры до ряда популяций Ирана и других соседних регионов (Табл.4).
Генетические расстояния мы вычисляли по (18), частоты генов талышей вычислены нами (Табл.1), а частоты генов сравниваемых с талышами популяций взяты из (2, 3,22,12). Как видно из Табл. 4, наименьшим является генетическое расстояние между талышами и иранцами Шираза (d= 0,23664), несколько дальше расположена от талышей популяция афганцев и иранцев Северо-Западного Ирана. Еще дальше расположены по генетическому расстоянию от талышей три популяции азербайджанцев, и иранцев Северного и Восточного Ирана и Тегерана. Столица Иранского Азербайджана Тебриз географически находится ближе к изучаемой нами популяции талышей, чем столица Азербайджана Баку. Генетическая близость некоторых популяций иранцев к талышам Азербайджана объяснима с учетом предшествующей истории этих популяций: возможно, в Талышские горы предки исследуемой нами популяции талышей пришли из региона Центрального Ирана. Приведенное для сравнения в Табл.4 генетическое расстояние талышей от русских Москвы значительно больше всех остальных генетических расстояний таблицы.
Таблица 4.
Генетические расстояния между популяцией талышей Пирасоры и популяциями соседних регионов.
|
Популяция, сравниваемая с талышами Пирасоры |
Генетическое расстояние |
Популяция, сравниваемая с талышами Пирасоры |
Генетическое расстояние |
|
Иранцы Шираза |
0,23664 |
Азербайджанцы Баш-Кунгута |
0,28262 |
|
Иранцы Северо-Западного Ирана |
0,27426 |
Азербайджанцы Джугута |
0,36662 |
|
Иранцы Северного Ирана |
0,30250 |
Азербайджанцы Сучмы |
0.28902 |
|
Иранцы Тегерана |
0,33284 |
Русские Москвы |
0,76706 |
|
Иранцы Восточного Ирана |
0,31444 |
||
|
Афганцы |
0,26051 |
Таким образом, генетические расстояния талышей от других соседних популяций, а также их языковая принадлежность к фарси, и особенности культуры позволяют считать талышей особым этносом, более близким к иранцам и афганцам, чем к азербайджанцам.
Кавказ представляет значительный интерес для популяционных антропогенетиков и антропологов своей особенной этнической пестротой. Среди исследователей, изучавших антропологический состав Кавказа, выделяется своими работами М.Абдушелишвили. Он исследовал 118 современных этно-территориальных групп Кавказа в краниологическом аспекте (23). Исследованию Кавказа посвящена также монография акад. В.П.Алексеева (24) Антропологическое изучение черепов, обнаруженных археологами в Душетском районе, датированных начиная с энеолита и до современности, было проведено одним из авторов данной работы В.О.Асланишвили. В развитом средневековье (Х-ХIV в.в.) было зафиксировано существование трех краниотипов: долихо- мезо и брахикранных. Население различалось и по формам лица, которых также выджелено три. Обнаружена также монголоидная и негроидная примесь. По мнению академика В.П.Алексеева, на территории Кавказа с древнейших времен существовали два европеоидных типа: брахикранный, широколицый, массивный, и долихокранный, узколицый, грацильный (24). В той же работе показано наличие монголоидной примеси на Северном Кавказе. М. Абдушелишвили, однако, не соглашался с наличием двух антропологических типов на территории Грузии, тем более с наличием каких либо не-европеоидных примесей. В работе В.В.Бунака (25) говорится о наличии 3-х краниотипов с преобладанием долихомезокранных над брахикранными. Повидимому, наряду с грузинскими племенами, в исследуемом районе Южного Кавказа проживало этнически различное население. В последующие эпохи постепенно происходила «грузинизация» этого населения, приведшая к формированию грузинского этноса. В табл. 5 приведены антропологические данные по популяции бассейна Арагви.
Наиболее ранний краниологический материал из Абаносхеви принадлежит к энеолиту
(IV тысячи лет до н.э.) Череп мужчины 25-30 лет резко долихокранный (64,8) с высоким переносьем (5,2 мм), симиотический указатель больше дакриального — череп с монголоидной примесью. Типология этого черепа кавказо-монголоидная. Наиболее близок этот материал к синхронными материалами из Армении (Шенгавит, Джарарит) и со среднеазиатскими черепами периода неолита (Овадантепе, Устпарим).
Черепа периода ранней бронзы (III тысячелетие до н.э.) из предгорной зоны (Квемо Араписи, Булачаури) — долихокранного типа, кавказо — европеоидные, из горной зоны (Георгиумидалирисской) — череп брахикранный, евро — африканский, а женские черепа долихо- и мезокранные — кавказо-африканские.
Таким образом, население этой части Южного Кавказа находилось в близком морфологическом сходстве с населением Грузии, Кавказа, Европы, Азии и Африки. Это не удивительно, поскольку на протяжении тысячелетий Кавказ был как бы узкими воротами из Азии в Европу, близко он расположен и к Африке. На западном побережье Каспийского моря, где Кавказские горы оставляют лишь узкую пролосу низменности, с древности проходил единственный удобный путь из степей Юго-Восточной Европы на Ближний Восток. Мигрирующие из Азии племена древних кочевников проходили по этому пути, иногда частично или полностью укрываясь надолго в горах и долинах Кавказа, где происходило как смешение племен, так и процессы, ведшие к их генетической изоляции. Ближайшие связи выявлены между краниологическими данными исследуемого региона с аналогичными материалами по баскам Испании и Франции.
Мы вычислили матрицу генетических расстояний 16 популяций Кавказа и Закавказья, а также соседних популяций зарубежных стран, в том числе СНГ, она приведена в Табл. 6.
Генетические расстояния вычисляли по 28 аллелям 12 локусов белков, ферментов и групп крови. Частоты генов популяций людей взяты нами из работ сотрудников нашей бывшей группы, из лаборатории генетики человека ИОГен РАН (7-10), а также из работ других отечественных и зарубежных авторов и нашей сводки (2-4, 22), частоты генов талышей и русских получены нами по собственным данным(Табл.1, 22).
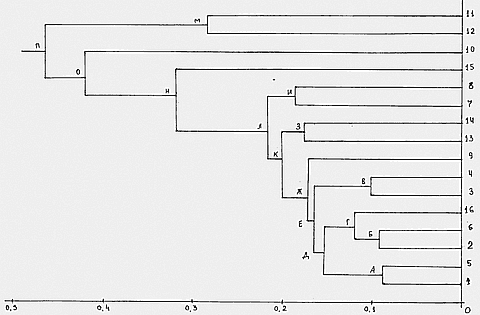
По значениям генетических расстояний из Табл. 6 мы построили эволюционную дендрограмму исследуемых популяций Кавказа, Закавказья и соседствующих регионов (Рис.1). Как видно на Рис. 1, исследуемые 16 популяций делятся на два кластера, в один из которых, значительно удаленный от популяций Кавказа и соседних регионов, входят монголы и негры Африки, как представители монголоидной и негроидной ветвей человечества (рас).
Остальные 14 популяций делятся на большой кластер, содержащий все популяции Кавказа и соседних регионов, и кластер, содержащий одну ветвь индийцев. Таким образом, население Кавказа по белкам, ферментам и группам крови ближе к европеоидам Азии — индийцам, чем к монголоидам или негроидам, что, впрочем, очевидно и по антропологическим признакам. Остальные популяции разделяются на соседние районы, существовала предковая популяция людей, которая начала дифференцироваться на отдельные популяции. Как видно из Рис. 2, раньше всех от общей популяции отделился
Таблица 5.
Краниологические и соматические материалы по популяции бассейна Арагви.
|
Ethnyc group Этническкая группа Type, Race Тип,Раса |
Khevsurs Хевсуры n=97 |
Gudamakrians Гудамакрианы n=109 |
Pshaves Пшавы n=105 |
Mtiuls Мтиулы n=102 |
Kakhies Кахетиныцы n=101 |
Mokheves Мохевы n=100 |
||||||
|
Index 8:1 |
n |
% |
n |
% |
n |
% |
n |
% |
n |
% |
n |
% |
|
Hiperbrachicephal 85.5 ® Гипербрахикефалы 85.5® |
16 |
16.50 |
40 |
36.69 |
64 |
60.95 |
32 |
31.37 |
40 |
39.60 |
32 |
32.00 |
|
Brachicephal 81.0 -85.4 Брахикефалы 81.0 -85.4 |
58 |
59.80 |
48 |
44.04 |
35 |
33.33 |
44 |
43.13 |
38 |
37.62 |
39 |
39.00 |
|
Mezocephal 76.0 — 80.9 Мезокефалы 76.0 — 80.9 |
23 |
23.70 |
21 |
19.27 |
5 |
4.76 |
22 |
21.56 |
20 |
19.82 |
27 |
27.00 |
|
Dolichocephal ® 75.9 Долихокефалы ® 75.9 |
– |
– |
– |
– |
1 |
0.95 |
4 |
3.92 |
3 |
2.97 |
2 |
2.00 |
|
Latitudo Nasalis |
||||||||||||
|
Caucasian — 33 Кавказцы — 33 |
8 |
8.24 |
12 |
11.02 |
15 |
14.70 |
7 |
6.86 |
22 |
21.78 |
10 |
10.00 |
|
European 34 — 36 Европейцы 34 — 36 |
34 |
35.05 |
32 |
29.35 |
35 |
34.31 |
38 |
37.25 |
39 |
38.61 |
34 |
34.00 |
|
Mongol 37 — 39 Монголы 37 — 39 |
37 |
38.14 |
46 |
42.20 |
37 |
36.27 |
39 |
38.23 |
26 |
25.74 |
39 |
39.00 |
|
Negro-African 40 — 50 Негры Африки 40 — 50 |
18 |
18.55 |
19 |
17.43 |
15 |
14.70 |
16 |
15.68 |
14 |
13.86 |
17 |
17.00 |
|
Crrassitudo labii 3, 4 |
||||||||||||
|
Negro-African Негры Африки |
8 |
8.25 |
6 |
5.50 |
5 |
4.76 |
8 |
7.84 |
10 |
9.90 |
10 |
10.00 |
Таблица 6.
Матрица генетических расстояний 16 популяций Кавказа и Закавказья и зарубежных стран
|
Популяции |
№п/п |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
15 |
|
Армяне |
1 |
|||||||||||||||
|
Грузины |
2 |
0.14437 ± 0.03660 |
||||||||||||||
|
Азербайджанцы |
3 |
0.12439 ± 0.03415 |
0.15154 ± 0.03742 |
|||||||||||||
|
Осетины |
4 |
0.18261 ± 0.04074 |
0.20258 ± 0.04267 |
0.09834 ± 0.03058 |
||||||||||||
|
Чеченцы |
5 |
0.08924 ± 0.02920 |
0.18122 ± 0.04060 |
0.13031 ± 0.03490 |
0.16099 ± 0.03847 |
|||||||||||
|
Талыши |
6 |
0.16452 ± 0.03886 |
0.09241 ± 0.02969 |
0.16543 ± 0.03895 |
0.14425 ± 0.03659 |
0.13585 ± 0.03558 |
||||||||||
|
Иранцы |
7 |
0.22267 ± 0.04448 |
0.17676 ± 0.04014 |
0.12595 ± 0.03435 |
0.19656 ± 0.04210 |
0.22953 ± 0.04508 |
0.23940 ± 004591 |
|||||||||
|
Арабы |
8 |
0.18659 ± 0.04113 |
0.17191 ± 0.03964 |
0.14835 ± 0.03706 |
0.21140 ± 0.04348 |
0.21296 ± 0.04362 |
0.10338 ± 0.03131 |
0.18730 ± 0.04120 |
||||||||
| Евреи Ближнего Востока |
9 |
0.16925 ± 0.03936 |
0.16889 ± 0.03932 |
0.12780 ± 0.03459 |
0.16889 ± 0.03932 |
0.06698 ± 0.03912 |
0.20200 ± 0.04261 |
0.21363 ± 0.04368 |
0.21803 ± 0.04408 |
|||||||
|
Индийцы |
10 |
0.32895 ± 0.05243 |
0.30496 ± 0.05084 |
0.31130 0.05127 |
0.36790 ± 0.05479 |
0.33862 ± 0.05304 |
0.31492 ± 0.05151 |
0.28053 ± 0.04911 |
0.30011 ± 0.05050 |
0.32318 ± 0.05205 |
||||||
|
Негры Африки |
11 |
0.42245 ± 0.05773 |
0.43023 ± 0.05811 |
0.39072 ± 0.05607 |
0.38318 ± 0.05566 |
0.39514 ± 0.05631 |
0.12505 ± 0.03424 |
0.49593 ± 0.06107 |
0.37340 ± 0.05511 |
0.46632 ± 0.05980 |
0.57587 ± 0.06404 |
|||||
|
Монголы |
12 |
0.32725 ± 0.05232 |
0.33084 ± 0.05255 |
0.27820 ± 0.04894 |
0.28593 ± 0.04950 |
0.33305 ± 0.05269 |
0.15988 ± 0.03835 |
0.32064 ± 0.05189 |
0.27931 ± 0.04902 |
0.27868 ± 0.04897 |
0.36683 ± 0.05473 |
0.28345 ± 0.04932 |
||||
|
Греки |
13 |
0.19312 ± 0.04177 |
0.22928 ± 0.04506 |
0.16380 ± 0.03878 |
0.17101 ± 0.03955 |
0.14212 ± 0.03634 |
0.13388 ± 0.03534 |
0.23859 ± 0.04584 |
0.24341 ± 0.04624 |
0.24622 ± 0.04647 |
0.37398 ± 0.05514 |
0.40732 ± 0.05695 |
0.34278 ± 0.05329 |
|||
|
Русские |
14 |
0.19159 ± 0.04229 |
0.22026 ± 0.04427 |
0.16426 ± 0.03883 |
0.16992 ± 0.03943 |
0.15131 ± 0.03740 |
0.18438 ± 0.04091 |
0.20538 ± 0.04293 |
0.23139 ± 0.04523 |
0.20761 ± 0.04313 |
0.39083 ± 0.05608 |
0.46859 ± 0.05990 |
0.38358 ± 0.05568 |
0.17502 ± 0.03996 |
||
|
Баски |
15 |
0.25761 ± 0.04737 |
0.18492 ± 0.04097 |
0.27706 ± 0.04885 |
0.32938 ± 0.05245 |
0.26125 ± 0.04776 |
0.15976 ± 0.03834 |
0.32671 ± 0.05228 |
0.34444 ± 0.05339 |
0.32059 ± 0.05188 |
0.45330 ± 0.05921 |
0.57781 ± 0.06410 |
0.50382 ± 0.06139 |
0.30210 ± 0.05064 |
0.24619 ± 0.04646 |
|
|
Испанцы |
16 |
0.14937 ± 0.03718 |
0.13119 ± 0.03501 |
0.15672 ± 0.03800 |
0.22827 ± 0.04497 |
0.15945 ± 0.03831 |
0.10957 ± 0.03218 |
0.24269 ± 0.04618 |
0.23566 ± 0.04568 |
0.24732 ± 0.04655 |
0.39449 ± 0.05627 |
0.45934 ± 0.05948 |
0.38110 ± 0.05554 |
0.19084 ± 0.04155 |
0.19662 ± 0.04211 |
0.18987 ± 0.04145 |
кластер, содержащий ветвь басков, и большой кластер, содержащий популяции Кавказа и Закавказья, а также ряд популяций Европы и Ближнего Востока. Далее кластер иранцев и арабов отделяется от кластера, содержащего остальные 11 популяций. Кластер, содержащий две ветви- русских и греков, отделяется далее, подтверждая данные Малярчука (26) о близости славянских популяций и популяций Южной Европы. Затем ветвь евреев Ближнего Востока отделяется от популяций Кавказа и испанцев. Популяции же непосредственно Кавказа и Закавказья разделяются следующим образом: сначала отделяется кластер, содержащий азербайджанцев и осетинов, затем ветвь испанцев отделяется от остальных этносов Кавказа и Закавказья,, и последние делятся на два кластера, содержещие один- грузин и талышей, и другой- армян и чеченцев. Испанцы имеют весьма смешанное происхождение, поскольку кельты, населявшие иберийский полуостров перед началом нашей эры, в первом тысячелетии нашей эры смешивались с германскими племенами вестготов, а во второй половине I тысячелетия н.э. большая часть Иберийского полуострова была занята арабами. Причем в Западно-Европейском эпосе « Песнь о Роланде » жившие в Испании арабы (мавры) именуются «испанцами». Смешение всех этих народов и дало испанскую нацию, причем кельты — баски до сих пор сохраняют свою национальную самоидентификацию. В первом тысячелетии нашей эры арабы неоднократно занимали часть Кавказа и Закавказья, ведя войны как с местными племенами, так и с хазарами (27, 28). Поэтому близость испанцев к народам Кавказа объяснима.
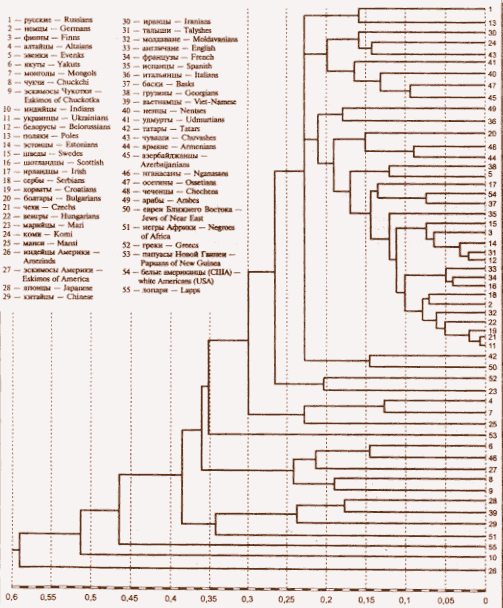
Ранее мы вычислили матрицу генетических расстояний 55 человеческих популяций, принадлежащих к по крайней мере четырем человеческим расам, и населяющим Европу, Азию, Америку, Африку и Океанию, по 28 аллелям 12 локусов белков, ферментов и групп крови (11). Все исследуемые в данной работе этнические популяции народов Кавказа также представлены в этой матрице. По этой матрице мы построили эволюционную дендрограмму 55 человеческих популяций, принадлежащих к 4 человеческим расам — европеоидов, монголоидов, негроидов и меланезийцев (Рис. 2). В соответствии с нашей гипотезой, опубликованной впервые в 1999 г. (12), и развитой в (30), в среднем палеолите (50-60 тыс. лет назад) в центре Азии, в большом регионе, включающем Алтай и азиатской прапопуляции отделились америнды — предки американских индейцев. Большой кластер, включающий 54 остальные популяции, ранее всего разделился на ветвь индийцев и кластер, содержащий все остальные популяции Из истории и археологии известно (31), что в III тысячелетии до н.э. населявшие полуостров Индостан племена дравидов, имеющих темную кожу и ряд других негроидных признаков, стали оттесняться к югу пришедшими из более северных регионов светлокожими европеоидными племенами. В дальнейшем проходило смешивание этих двух популяций, в результате чего и образовались народности, населяющие Индию. Поэтому, видимо, индийцы и представляют на дендрограмме (Рис.2) ветвь, ранее отделившуюся от остальных популяций Евразии.
Далее от оставшихся популяций отделилась ветвь лопарей (саамов). Как показано нами в (12, 30) лопари были, повидимому, первыми древними европеоидами, заселившими Север Европы. Обнаружение археологических культур верхнего палеолита в Финляндии (32) также указывает на очень раннее заселение Северной Европы.
Затем из дендрограммы (Рис. 2) следует, что от огромного кластера европеоидных и североазиатских популяций отделился кластер, содержащий ветвь негроидов, ранее отделившуюся от кластера монголоидов Юго-Восточной Азии — китайцев, японцев и вьетнамцев.
Из кластера европеоидов и монголоидов Северной Азии прежде всего выделился субкластер, содержащий популяции якутов, нганасан, американских эскимосов, чукчей и азиатских эскимосов.
Как показано нами в работе (12), дифференциация всех перечисленных выше популяций происходила, повидимому, на территории глубинной Азии со среднего палеолита до неолита, и сопровождалась длительной миграцией популяций от места их дифференциации до современных мест обитания. На основании обнаруженной нами по частотам аллелей полиморфных локусов белков, ферментов и групп крови европеоидной компоненты алтайцев (33), а также обнаруженной другими авторами идентичности гаплогрупп митохондриальной ДНК у алтайцев и американских индейцев (34), мы предположили, что популяции северных монголоидов, америндов и ряда европеоидных народов происходят из одной предковой популяции, обитавшей в центре Азии во времена среднего палеолита (12, 30).
Возвращаясь к Рис.2, мы видим, что далее от кластера, содержащего остальные 42 популяции обитателей Евразии, отделилась ветвь папуасов Новой Гвинеи. По антропологическим данным А.А.Зубова (35), папуасы отделились от остального человечества после отделения америндов, что подтверждает результаты нашей дендрограммы.
Следующим из кластера, содержащего остальные 42 популяции, выделился субкластер, содержащий ветви, соответствующие популяциям алтайцев, монголов и манси (Рис.2). В отдельный субкластер объединены популяции греков и марийцев. Это на первый взгляд непонятное объединение можно объяснить тем, что марийцы, являющиеся уже настоящими европеоидами, могут быть близки к древним европеоидам, еще за несколько тысяч лет до н.э. мигрировавшим в Европу, скорее всего, из Азии, — ахейцам, представителям Крито-Микенской цивилизации, потомками которой являются греки.
Татары и евреи Ближнего Востока образуют две ветви кластера, также рано отделившегося от остальных популяций, что можно объяснить общим происхождением татар и евреев; значительная часть последних является мигрантами из Восточной Европы, а до того из тюркских азиатских популяций: известно, что в VIII-X в.в. н.э. евреи жили в одном государстве, Хазарском каганате, с тюрками-хазарами, и после гибели в X в. этого государства мигрировали с Нижней Волги в более западные районы Восточной Европы (27, 28). Ранее жители Хазарии вытеснили с Нижней Волги и Прикавказских степей древних булгар в более северные районы Поволжья и Прикамья (27), в результате чего обитающие там в настоящее время татары несут в себе значительную древнебулгарскую компоненту (28). Во время этих процессов могла иметь место метисация древних булгар с жителями Хазарии, часть которых была этническими евреями. Однако большая часть жителей Хазарии была тюрками, принявшими иудаизм; часть евреев Восточной Европы является их потомками.
Остальные 35 популяций европеоидов дифференцировались следующим образом: они разделились на два кластера, к одному из которых относится субкластер русских и поляков, а также два субкластера, один из которых содержит ветвь иранцев, ветвь коми и ветвь чувашей, а другой — ветви удмуртов, ненцев, осетинов и азербайджанцев (Рис. 2). Как показано нами в предыдущих работах (12, 30), ненцы представляют как бы след миграции древних праевропеоидных популяций с мест их первичной дифференциации в центре Азии через Север Сибири на Север Европы. Осетины являются потомками аланов — скифских европеоидных древнеиранских племен (28). Дифференциация этой части большого кластера европеоидных популяций иллюстрирует этногенез славян, финно-угров и иранцев, происходивший, вероятнее всего, в конце верхнего палеолита — неолите.
Некоторые историки считают, что русские, украинцы и ряд других славянских народов происходят от древнеиранских скифских племен, мигрировавших ранее по Азии (37). Геродот (36) именовал племена, обитавшие в его время на территории Восточной Европы, « скифами- пахарями». Американский археолог Мария Гимбутас считала, что индоевропейские племена мигрировали в Европу из Азии (38). М.В. Ломоносов в своих «Записках по русской истории» (39) писал о «древнем обитании славян-вендов в Азии».
Ближе всех, по данным дендрограммы (Рис.2), к русским находятся поляки. По антропологическим данным акад. Т.И. Алексеевой, славяне очень неоднородны по краниометрическим и другим морфологическим признакам (13,14). По ее мнению, среди славянских народов существует пять антропологических типов, образующих компактные ареалы. Беломоро-Балтийский тип (термин Н.Н. Чебоксарова) представляют белорусы, поляки и северные группы русских. Восточноевропейский тип (назван И.Е. Деникером) — это остальные территориальные группы русских, часть белорусов. В Днепро-Карпатский тип славян (В.В. Бунак) включают украинцев, словаков, часть чехов, прикарпатские этнические группы. По нашей дендрограмме (Рис. 2), в один субкластер входят украинцы и чехи, рядом находятся ветви венгров и молдаван.
В соседний кластер входит субкластер арабов и итальянцев. Малярчук также показал близость восточных славян (русских) к популяциям Южной Европы по маркерам митохондриальной ДНК (26). Этим субкластером начинается дифференциация большого кластера европеоидных популяций Европы и Азии, содержащего 26 народностей. После отделения арабов и итальянцев на Рис.2 видно, как идет разделение остальных 24 популяций этого кластера: отделяется субкластер, содержащий ветвь болгар, и две ветви — чеченцев и армян. Повидимому, три последние популяции происходят от древних европеоидных популяций, мигрировавших на просторах Передней Азии, большого региона Кавказа, Закавказья и Причерноморья. Выше мы упоминали о миграции части древних булгар в I тысячелетии н.э. из Западного Предкавказья, где в результате распада Западно-Тюркского Каганата в VII в. образовалась Великая Булгария (28), в регион Поволжья и Прикамья. В конце VII в. Великая Булгария потерпела поражение от хазар, и часть булгар с ханом Аспарухом мигрировала на Нижний Дунай, образовав там Дунайскую Булгарию, в населении которой в последующие века смешивалось местное славянское население и потомки тюрков-булгар.
На первый взгляд кажется странным нахождение в одном субкластере эвенков и грузин (Рис 2), но мы еще в 1984 году показали наличие у эвенков европеоидной компоненты по частотам аллелей белков, ферментов и групп крови (6). В работе (12) мы показали, что эвенки являются как бы следом миграции древних европеоидов из регионов глубинной Азии через Сибирь и Циркумполярную зону.
Далее на Рис. 2 видно выделение субкластера, содержащего популяции ирландцев, а также белых американцев и басков; по-видимому, эти популяции объединяет кельтский антропологический субстрат, присутствующий в них.
Большой кластер, объединяющий германские, финно-угорские и другие популяции Европы, подразделяется следующим образом: сначала отделяется ветвь испанцев — этот этнос, кроме указанных выше этнических компонентов, несет в себе генетический материал вестготов. После этого отделяются два субкластера, один из которых содержит ветви шведов, финнов, эстонцев, талышей и белорусов. Большинство из этих популяций обитают ныне в Прибалтийском регионе или вблизи него. Близость талышей Закавказья к данным популяциям может указывать на возможный путь миграции этой части европеоидов с их азиатской прародины.
Кельтский субстрат, видимо, объединяет в один субкластер ветви англичан, французов, шотландцев: англичане содержат генофонд кельтов — англов, французы — галлов и шотландцы — гэлов (Рис.2). В последний субкластер европеоидов входят ветви сербов, немцев, молдаван, венгров, хорватов, чехов и украинцев — популяции Центральной и Восточной Европы, предки которых, по нашему мнению, из Передней Азии, Причерноморья, а ранее из районов глубинной Азии (например, предки венгров — хунну) мигрировали по азиатским и южнорусским степям в Европу. В антропологических исследованиях Т.И. Алексеевой (13,14) обнаружено, что в некоторых группах западных средневековых славян по краниометрическим признакам прослеживаются черты, типичные для германцев.
Популяции Кавказа и Закавказья на большой де ндрограмме кластеризуются следующим образом: субкластер, содержащий ветви осетин и азербайджанцев, отделяется от ветви иранцев только четырьмя популяциями (Рис.2). Осетины — это потомки аланов, древнеиранских скифских племен. Нахождение в одном субкластере (Рис.2), как и на малой дендрограмме (Рис.1) ветвей армян и чеченцев объяснимо, так как сейчас стало известно об общности хуррито-урартской и вайнахской языковых семей (. Происхождение чеченцев мало изучено, есть неясные сведения о вхождении некоторых арабских родов в популяцию предков чеченцев, и действительно, ветвь арабов, входящая в один кластер с итальянцами, отделяется от субкластера армян и чеченцев только ветвью болгар. Выше мы писали о существовании Булгарского каганата во второй половине тысячелетия нашей эры в регионе степей Северного Кавказа. Армяне — потомки европеоидов Передней Азии, в древности в регионе теперешней Армении существовало государство Урарту. Т.И.Алексеева, В.П.Алексеев и М.Г.Абдушелишвили пишут о большом ареале физического типа динарцев, простирашимся от Балкан до Кавказа — этоантропологический тип массивных, широколицых людей с сильным выступанием носа (14, 24,25). В целом эти популяции Кавказа и Закавказья являются европеоидами. Однако вхождение популяции грузин в один субкластер с эвнеками (Рис.2), также, как и обнаружение монголоидной примеси в черепах из Грузии (Табл. 5) позволяет предположить, что мигрировавшие в палеолите по Азии популяции древних европеоидов, имевших еще и монголоидные признаки, могли в дальнейшем мигрировать в Кавказский регион.
СПИСОК ЛИТЕРАТУРЫ
1. Семенская Е.Л. Изучение групп крови народов Кавказа. // Советская Этнография. 1936.- № 4.-С.213.
2. Краснопольская К.Д., Филиппов И.К., Сотников Е.И. и др. Закономерности распределения аллелей Gd- в Азербайджане. I. Частота и полиморфизм на недостаточность глюкозо-6-фосфатдегидрогеназы в Шекинском районе Азербайджана. // Генетика.- 1980.-9, № 9.-С. 1693-1700.
3. Мовсум-Заде К.М., Расулов Э.М., Аскерова Т.А. Исследования генетической гетерогенности Г-6-ФД недостаточности в Азербайджанской СССР. I. Данные по структуре, распределению некоторых групп крови и антропологических признаков в трех деревнях Массалинского района. //Изв. Ан Аз. ССР, сер. биол.-1980.-99, № 5.
4. Булаева К.Б., Павлова Т.А., Булаев О.А. Генетический полиморфизм в трех популяциях коренных народов Дагестана.// Генетика.- 1997.- 33, №10.-С.1395-1405.
5. Nazarova A.F. The genetic structure of populations of Chuckotka peninsula Eskimos and Chuchchi based on the study of 13 loci of serum and erythrocyte proteins and enzymes. // Amer. J. Phys. Anthropol.- 1989.- 79, № 1.- P.81-88.
6. Рычков Ю.Г., Спицын В.А., Шнейдер Ю.В., Назарова А.Ф. и др. Генетика популяций таежных охотников-оленеводов Средней Сибири. Биохимические маркеры генов HP, Tf, Gc, Alb, GLO I, PGM I, AcP и Es D. // Генетика.- 1984.- 20, № 10.-С. 1701-1707.
7. Инсаридзе З.П., Насидзе И.С., Шенгелия Л.А. и др. Генетика народонаселения Кавказа: распределение некоторых иммунологических и биохимических маркеров в Западной Грузии. // Генетика.- 1990.- 26, № 6.- С. 1092-1101.
8. Инсаридзе З.П., Насидзе И.С., Шенгелия Л.А. и др. Генетика этнических групп Кавказа: распределение некоторых иммунологических и биохимических маркеров в Северной Осетии и Чечено-Ингушетиии. // Генетика.- 1990.- 26,№ 9.- 1648-1659.
9. Насидзе И.С., Инсаридзе З.П., Шенгелия Л.А. и др. Генетика народонаселения Кавказа: распределение некоторых иммунологических и биохимических маркеров в Восточной Грузии. // Генетика.- 1990.- 26, № 5.- С.936-945.
10. Шенгелия Л.А., Инсаридзе З.П., Насидзее И.С. и др. Генетика народонаселения Кавказа: распределение некоторых иммунологических и биохимических маркеров в азербайджанской и армянской популяциях в Грузинской ССР. // Генетика.- 1991.-27, № 10.- С. 1832- 1839.
11. Назарова А.Ф., Алхутов С.М. Генетические расстояния 55 человеческих популяций Европы, Азии, Америки, Африки и Океании по 28 аллелям 12 локусов белков, ферментов и групп крови.// Эволюция человеческих популяций.- М.: Лип. изд-во, 2000.
12. Назарова А.Ф. Популяции, переходные между монголоидами и европеоидами, и возможный путь формирования европеоидов. // Генетический портрет народов мира.- М.: Лип. из д-во, 1999.-С.4-16.
13. Алексеева Т.И. // Этногенез восточных славян по данным антропологии.- М.: Изд-во МГУ, 1973.- 329 С.
14. Алексеева Т.И. Этногенез и этническая история восточных славян. // Восточные славяне. Антропология и этническая история. — М.: Научный мир, 1999.- С. 307-315.
15. Gahne B., Junea R.K., Grolmus J. Horizontal polyacrilamide gradient gel electrophoresis for the simultaneous phenotyping of transferrin, posttransferrin, albumin and postalbumin in the blood plasma of cattle. // Anim. Blood Gr. Biochem. Genet. — 1977.-8.- P. 127-137.
16. Nei M., Roychoudhury A.K. Sampling variances of heterozygosity and genetic distance. // Genetics.- 1974.-76.-P.379-390.
17. Selander H. Behaviour and genetic variation in natural populations. // Amer. Zoologist.- 1970.-10.-P. 53-66.
18. Cavalli-Sforza L.L., Bodmer W.F. // The genetics of human populations. — San Francisco: WH Freeman Co.- 1971.
19. Машуров А.М., Черкащенко В.И. Учитывать генетические дистанции между породамию// Живот новодство.- 1987.-№2.- С.21-23.
20. Назарова А.Ф. Генетическая структура паопуляции талышей Азербайджанской ССР.// Доклады РАН.- 1991.- 317,№6.- С.1484-!486.
21. Раутиан Г.С., Ата-Мурадова Ф.А., Мироненко Е.В. Миграционная структура и инбридинг в популяции талышей. // Генетика.- 1993.- 29.- С.1035-1041.
22. Назарова А.Ф., Алхутов С.М. // Генетический портрет народов мира. М.: Лип. изд-во.- 1999.-32 С.
23. Абдушелишвили М. Г. // Антропология древнего и современного населения Грузии.- Тбилиси.- Мецниереба.-1964.
24. Алексеев В.П. // Происхождение народов Кавказа. — М.- Наука, 1974.- 315 С.
25. Бунак В.В. Антропологический состав населения Кавказа.// Вестник Гос. музея Грузии.- Xii-A, 1946.
26. Малярчук Б.А. Распределение маркеров смитохондриальной ДНК в европеоидной популяции Евразии // Генетика.- 1997.-33, № 7.- С.986-991.
27. Артамонов М.И. // История хазар. -Л.: Наука, 1962.
28. Новосельцев А.П. // Хазарское государство и его роль в истории Восточной Европы и Кавказа.-М.: Наука, 1990.
29. Назарова А.Ф., Алхутов С.М., Машуров А.М. // Эволюция человеческих популяций. М.: Лип. изд-во, 2000.
30. Назарова А.Ф. К проблеме дифференциации северных монголоидов, европеоидов и америндов на территории Евразии (генетические данные) // Цитология и генетика.-2002.- 36, № 6.- С.46-53. 31. Бонгард-Левин Г.М., Масон В.М. и др. // Археология зарубежной Азии.-М.: Наука, 1986.
32. Кетола П. // Белое пятно.- Ketola P.// Valkoinen Paikka. Helsinki,1966.
33. Назарова А.Ф., Кузнецова М.Г. Генетическая структура популяций алтайцев // Доклады РАН.- 1993.-333, № 3.- С. 405-409.
34. Сукерник Р.И., Шур Т.Г., Стариковская Е.Б. и др. Изменчивость митохондриальных ДНК коренных жителей Сибири в связи с реконструкцией эволюционной истории американских индейцев // Генетика- 1996.- 32, № 3.- С. 432-439.
35. Зубов А.А. Проблема гомогенности физического типа коренного населения Америки // Тез.докл. III Конгресса этнографов и антропологов России. — М, 1999.- С.107.
36. Геродот // История.
37. Гобарев В.М. // Предыстория Руси.- М.: Менеджер, 1994.- Ч.1.- 331 С.
38. Gimbutas M. Old Europe in the Fifth millenium B.C.: the European situation on the arrival of indo-europeans // The Indo-Europeans in the Fourth and Third millenia. Ann Arbo.- 1982, P.34
39. Ломоносов М.В. // Записки по русской истории.- М.- ЭКСМО.- 2003.- 704 С.
40. Diakonov I.M., Starostin S.A. Hurrito-Urartian as a Eastern Caucasian Language. 1987, Munchen.
РЕЗЮМЕ
Проведенные нами антропологическик исследования показали, что в бассейне реки Арагви начиная с энеолита жили представители разных человеческих рас, а именно европеоидной и монголоидной. Эти данные подтверждают мнение академика В.П. Алексеева (1974) о смешанном в расовом отношении населении Кавказа с древних времен.
Исследовали электрофоретический полиморфизм белков крови популяций талышей Пирасоры, расположенной в Талышских горах на Юго-Востоке Азербайджана. Вычислив частоты генов популяции талышей, мы определили генетические расстояния талышей до иранцев Северного, Центрального и Южного Ирана, афганцев и трех популяций азербайджанцев. Оказалось, что талыши наиболее близки к иранцам Шираза, а популяции азербайджанцев наиболее удалены от талышей. Проведенные нами антропологическик исследования показали, что в бассейне реки Арагви начиная с энеолита жили представители разных человеческих рас, а именно европеоидной и монголоидной. Эти данные подтверждают мнение академика В.П. Алексеева (1974) о смешанном в расовом отношении населении Кавказа с древних времен. Мы вычислили матрицу генетических расстояний ряда народов Кавказа и Закавказья, в том числе грузин, осетин, азербайджанцев, армян и других до этносов соседствующих регионов и до басков по 28 аллелям 12 локусов белков, ферментов и групп крови, и построили дендрограмму этих популяций. По ранее вычисленной нами матрице генетических расстояний 55 человеческих популяций, принадлежащих к четырем расам, мы построили дендрограмму. Положение народов Кавказа на обеих дендрограммах в основном соответствует данным отечественных антропологов. Кластеризация других европеоидных этносов в целом также соответствует антропологическим и историческим данным, и подтверждает выдвинутую нами ранее гипотезу (Назарова, 1999) о дифференциации европеоидов, северных монголоидов и американских индейцев из одной популяции, обитавшей в палеолите в Азии.
Адрес для переписки с авторами:
Назаровой Ариадне Филипповне.
E.mail: afnazar@yandex.ru
 |
|
Рис.1. Дендрограмма 16 популяций Кавказа и соседних регионов и некоторых других популяций. На горизонтальной оси отложены значения генетических расстояний. |
|
|
1 -Армяне |
9 — Евреи Ближнего Востока |
|
2 -Грузины |
10 — Индийцы |
|
3 Азербайджанцы |
11 -Негры Африки |
|
4 -Осетины |
12 — Монголы |
|
5 — Чеченцы |
13 — Греки |
|
6 — Талыши |
14 — Русские |
|
7 — Иранцы |
15 — Баски |
|
8 -Арабы |
16 -Испанцы |
 |